当前位置:首页> >肠动力研究院> >技术文章> > 技术文章
第299期:T-2毒素通过促进IL-6/JAK/STAT1介导的炎症和凋亡破坏肠道菌群-脾脏轴诱导的脾脏损伤及纳米硒的
论文ID
原名:T-2 toxin-induced splenic injury by disrupting the gut microbiota - spleen axis via promoting IL-6/JAK/STAT1 signaling-mediated inflammation and apoptosis and its mitigation by elemental nano‑selenium
译名:T-2毒素通过促进IL-6/JAK/STAT 1信号介导的炎症和凋亡破坏肠道菌群-脾轴诱导的脾损伤及元素纳米硒的缓解作用
作者:Meng Liu1, Xue Wu Li1,2, Hua Sun3, Yi Qin Yan1, Zhi Yuan Xia1, Alainaa Refaie1, Ni‑Ya Zhang1, Shuai Wang1, Chen Tan1, Lv‑Hui Sun1
完成单位:
期刊:Archives of Toxicology
影响因子:4.8
发表时间:2025-02-27

1、摘要
T-2毒素是环境和食物链中普遍存在的毒性最大的单端孢菌毒素之一,给动物和人类带来严重的健康危害,并能显著损害免疫功能。本研究通过整合转录组和微生物组,全面解释了T-2毒素通过肠道微生物-脾脏轴对脾脏的影响。结果表明,日粮T-2毒素≥1.0 mg/kg显著抑制肉仔鸡的生长性能,引起脾脏损伤,并伴有氧化应激和组织病理学损伤。盲肠微生物组分析表明,T-2毒素暴露导致肠道微生物生态失调,特别是导致一些增强肠道屏障和减少炎症的有益菌属减少。转录组分析表明,T-2毒素暴露直接引起脾脏炎症和免疫相关信号,如细胞因子-细胞因子受体相互作用、Toll样受体信号通路、NOD样受体信号通路和JAK-STAT信号通路。
此外,通过联合转录组和微生物组分析,发现T-2毒素诱导的脾脏损伤与IL-6/JAK/STAT1信号通路介导的炎症和凋亡的异常激活有关,并通过western bolt分析验证。值得注意的是,日粮硒补充剂可以通过抑制IL-6/JAK/ STAT1信号相关基因的表达来保护雏鸡免受T-2毒素诱导的对生长性能和脾脏损伤的不利影响。总之,本研究为T-2毒素在鸡脾脏中的免疫毒性机制提供了新的见解,并强调了硒在减轻T-2毒素诱导的免疫毒性方面的潜力。
2、试验设计

3、结果与分析
3.1 T-2毒素降低生长性能并诱导脾损伤
4组肉鸡的初始体重没有差异(图1A)。经过2周的试验处理后,添加1.0mg T-2毒素/kg的日粮有降低(P=0.06)体重(12.4%)的趋势,添加3.0和6.0mg/kg的日粮有降低(P<0.05)最终体重(17.6–27.2%)的趋势(图1A)。在不同剂量下,饮食中T-2毒素补充≥1.0 mg/ kg可使体重增加减少13.7–30.2%(P< 0.05)(图1B)。此外,组织学结果显示,日粮补充T-2毒素诱导免疫组织损伤(图1C)。具体而言,与对照组相比,日粮添加T-2毒素≥1.0 mg/kg诱导脾淋巴细胞减少、网状上皮细胞增生和轻度充血,尤其是添加6.0 mg/kg T-2毒素的日粮诱导严重充血和巨噬细胞增生。同时,T-2毒素引起淋巴细胞坏死和耗竭、局灶性坏死、皮质淋巴细胞增殖、胸腺血管充血和法氏囊完整性的丧失。此外,T-2毒素诱导脾脏氧化损伤,与对照组相比,随着日粮T-2毒素剂量的增加,MDA和PC浓度分别增加55.7-97.3%和182.0-393.1%(P<0.05),T-AOC分别减少24.3-35.9%(P<0.05),呈剂量依赖性(图1D-F)。
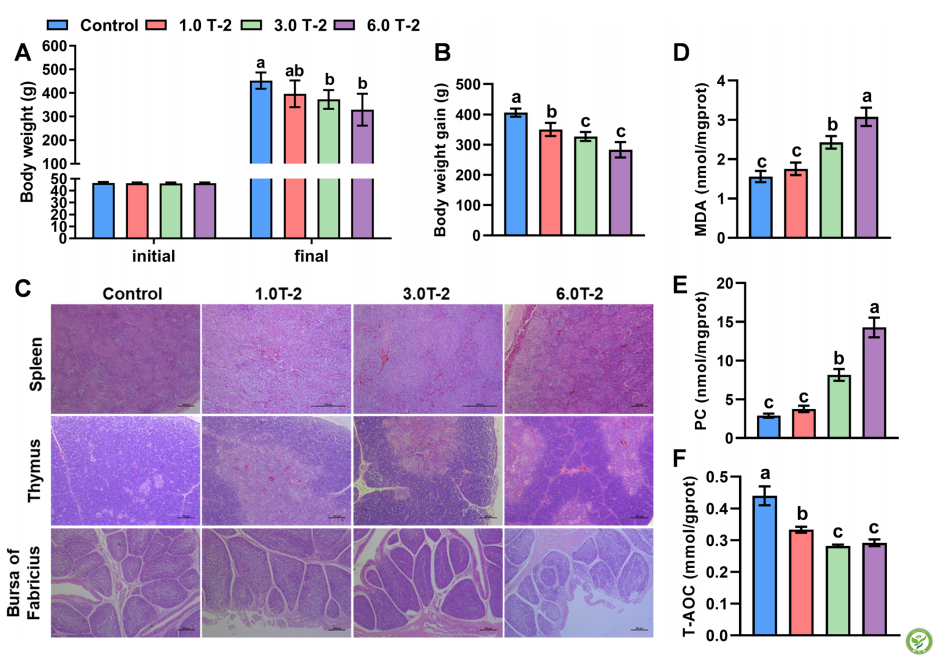
3.2 T-2毒素对盲肠菌群稳态的影响
为更好地了解T-2毒素对盲肠细菌组成的影响,对对照组和实验1中6.0 mg/kg T-2毒素组肉鸡进行了16 S rRNA微生物基因的高通量测序(图2)。根据序列相似性(>97%)将符合条件的序列聚类为OTU,T-2毒素处理组中OTU数量较少,两组共有499个OTU(图2A)。基于分类学组成分析,T-2毒素改变了盲肠细菌在门和属水平上的相对丰度。所有来自盲肠食糜样品的合格序列归属于7个已知门。在所有样品中,以厚壁菌门、拟杆菌门、变形菌门和软壁菌门为优势菌门(图2B)。与对照组相比,添加6.0mg/kg的T-2毒素可显著降低厚壁菌门和软壁菌门的丰度,提高拟杆菌门和变形菌门的丰度;并从所有合格序列中鉴定出50个优势属,包括拟杆菌属、嗜双歧杆菌属、乳杆菌属、粪大肠杆菌属和示波螺旋菌属(图2C)。微生物区系的α多样性通过Chao 1分析评估物种丰富度,Shannon指数评估物种均匀度。与对照组相比,在日粮中添加6.0 mg T-2毒素/ kg不会影响Chao 1指数(图2D),但会降低Shannon指数(图2E)。为确定两组之间不同的细菌分类群,使用了LDA>3的LEfSe分析。结果在对照组和T-2毒素组的微生物群中分别鉴定了8个和2个区别性特征(图2F)。此外,菌属的丰度显示,与对照组相比,T-2毒素组中的Blautia、Coprococcus、Lachnospira、Coprobacillus、Anaerostipes 属的丰富度显著减少;然而,与对照组相比,在T-2毒素处理下,Fusibacter丰度显著增加(图2L)。
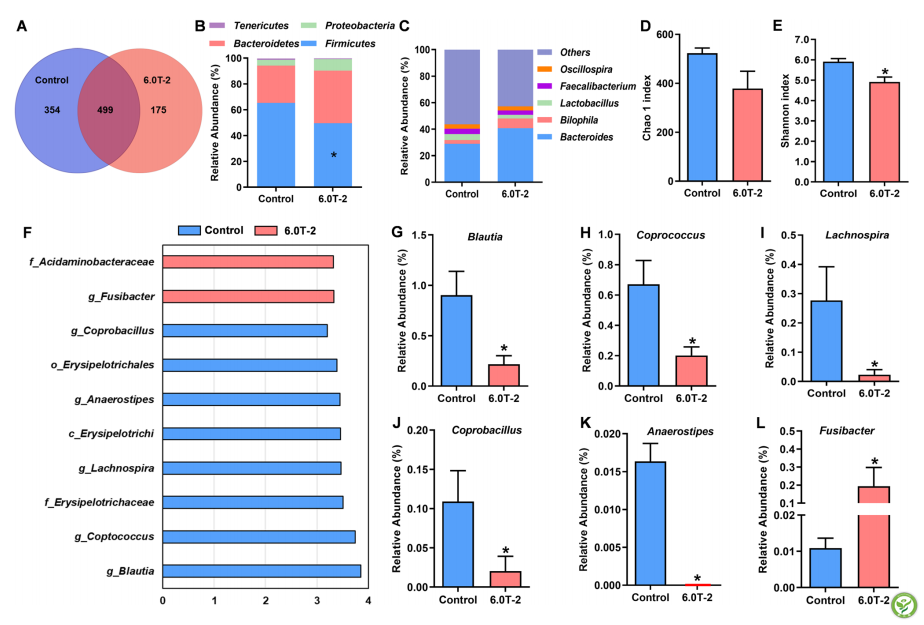
图2:日粮T-2毒素对盲肠微生物组的影响。A对照组和6.0 T-2组中OTU的数量。B门水平的肠道细菌分布。C属水平的肠道细菌分布。D和E T-2毒素对Chao 1指数的影响。E T-2毒素对Shannon指数的影响。F线性判别分析(LDA)效应大小(LEfSe)微生物群组成分析。G-L Blautia、Coprococcus、Lachnospira、Coprobacillus、Anaerostipes 、Fusibacter相对丰度。对照,基础饮食;6.0 T-2,基础日粮添加6.0 mg/kg T-2毒素。
3.3 T-2毒素对脾脏转录组的影响
为进一步揭示T-2毒素的毒性机制,对实验1的肉鸡脾脏组织进行了转录组分析。差异表达(DE)分析结果表明,T-2毒素处理的肉鸡脾脏中存在T-2毒素处理应答基因(图3)。与对照组相比,在日粮中补充1.0、3.0和6.0mg/kgT-2毒素时,分别有2489、1943和3936个基因表现出1.5倍或更高(P<0.05)的DE(图3A-C)。值得注意的是,在不同的T-2毒素剂量下,有907个基因具有相似的变化(图3D)。此外,GO和KEGG通路分析确定了共表达的DEG。主要GO类别的不同分布如图3E所示。在生物过程(BP)类别中,丰富了细胞周期、防御反应、免疫系统过程、对生物刺激的反应和对应激的反应。在细胞化合物(CC)类别中,染色质、染色体、胞外区细胞外空间和超分子复合物是GO富集。CCR趋化因子受体结合、CCR 6趋化因子受体结合、趋化因子活性、趋化因子受体结合和蛋白质异二聚体活性也是GO表达最多的类别。KEGG富集结果表明,KEGG的主要通路包括细胞因子-细胞因子受体相互作用、Toll样受体信号通路、核苷酸寡聚化结构域(NOD)样受体信号通路、IL-17信号通路、细胞坏死、凋亡-多物种、细胞因子生长因子和Janus激酶(JAK)-信号传导子和转录激活子(STAT)信号通路(图3F)。
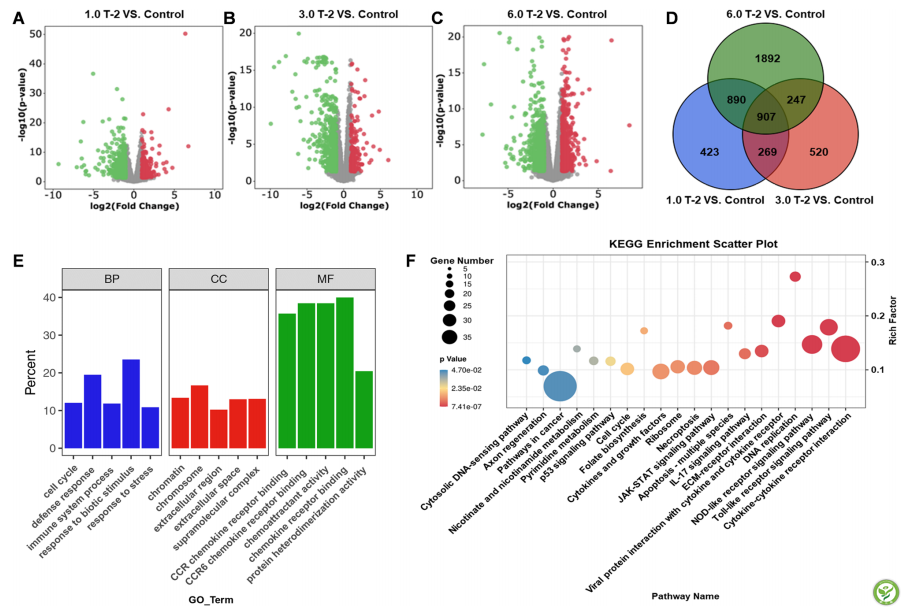
图3 :T-2毒素对脾脏转录组的影响。A-C火山图分别显示1.0 T-2、3.0 T-2、6.0 T-2毒素与对照中的DEG。D维恩图分析1.0 T-2毒素与对照、3.0 T-2毒素与对照、6.0 T-2毒素相对于对照DEG数量。E GO功能富集分析。F KEGG途径富集分析,对照,基础饲料; 1.0 T-2、3.0 T-2和6.0 T-2,基础饲料添加1.0、3.0和6.0 mg/kg T-2毒素。
3.4 转录组和微生物组联合分析揭示T-2毒素对脾脏的毒性机制
DEG与最丰富的细菌属的相关性分析表明,有17个DEG与10个属的DEG具有显著的相关性(图4A)。其中,Faecalibacterium_prausnitzii和Ruminococcus与大多数基因相互作用最高。此外,在KEGG途径分析中具有显著相关性的这些DEG的注释途径主要与炎症反应和免疫调节有关,包括ECM-受体相互作用、JAK-STAT信号传导途径和细胞因子-细胞因子受体相互作用(图4B)。基于筛选的KEGG富集途径,利用Western blot分析JAK-STAT信号通路在T-2毒素对肉鸡脾脏的影响中的作用(图4C和D)。值得注意的是,与对照相比,日粮补充1.0、3.0和6.0 mg T-2毒素/kg均增加了(P<0.05)JAK 2、JAK 3和c-Jun-N-末端激酶(JNK)的蛋白生成。对照组相比,3.0和6.0 mg T-2毒素/kg组显著增加(P<0.05)STAT 1、IL-6和p53蛋白的表达。有趣的是,6.0 mg T-2毒素/kg组的脾中p-STAT 1表达比其他三组低。然而,BCL-2的蛋白质产生不受日粮T-2毒素剂量的影响(P>0.05)(图4C和D)。
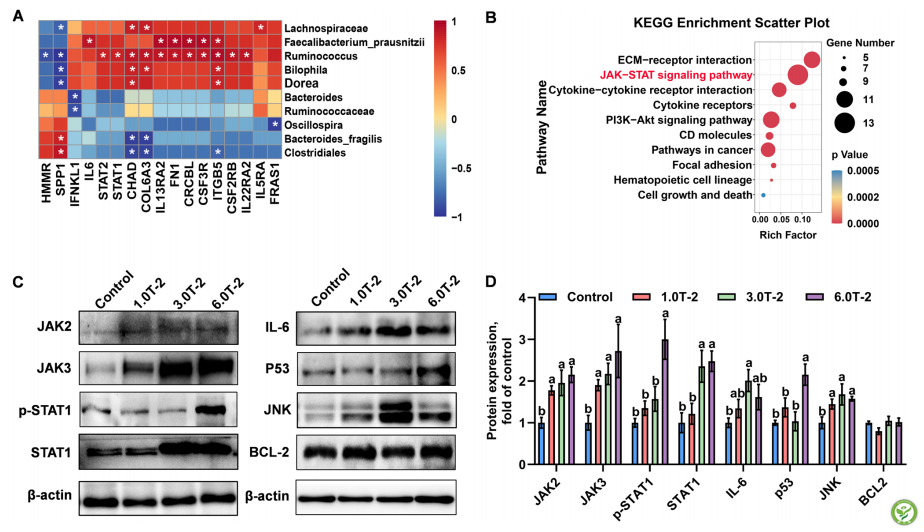
图4:转录组与微生物组的联合分析。A属与DEG之间的Pearson相关性分析。B KEGG途径富集分析DEG与属显著相关性。C参与JAK-STAT信号通路的蛋白相对丰度。D参与JAK-STAT信号通路的蛋白的相对丰度。
3.5 硒和T-2毒素对生长性能、脾脏氧化还原状态和组织病理学的影响
四组肉鸡的初始体重差异不显著(图5A)。在3周的实验处理后,与BD组相比,日粮T-2毒素补充降低了(P<0.05)终末体重(20.1%),体增重(21.9%),采食量(13.7%)和增重/饲料(9.2%),而补硒(添加硒为单质纳米硒,由北京德元顺生物科技有限公司提供)减轻了T-2毒素引起的这些变化(图5B-E)。与BD组相比,BD +T-2组增加了(P<0.05)GPX活性,BD+Se+T-2组降低(P<0.05)MDA含量。且与BD+T-2组相比,BD+Se+T-2组GPX活性升高(P<0.05)。然而,TrxR活性不受日粮Se缺乏和T-2毒素补充的显著影响(图5H)。除GPX活性外,没有观察到Se和T-2毒素之间的相互作用。此外,脾组织学结果显示,日粮T-2毒素诱导的脾损伤,这是由淋巴细胞数量减少和组织细胞增殖的减少所表明(图5I)。单独硒缺乏也可引起脾淋巴细胞减少和组织细胞增殖能力下降,而BD+Se组未见明显病理改变。
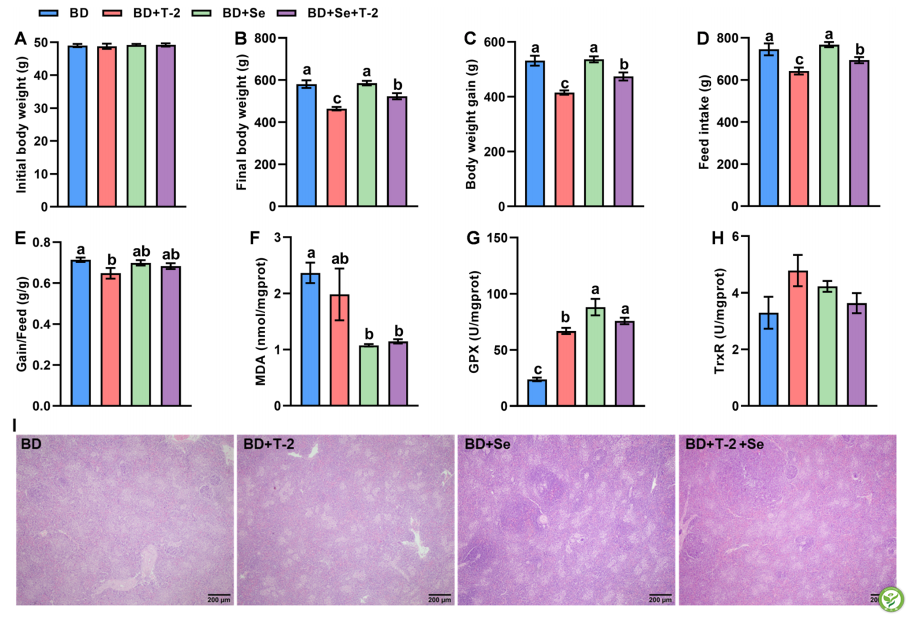
图5:日粮硒水平和T-2毒素对生长性能(A-E)、脾脏氧化还原状态(F-H)和组织病理学(I)的影响。BD:基础日粮; BD+T-2:BD +1.0 mg T-2毒素/kg; BD+Se,BD +0.2 mg Se/kg; BD+T-2+Se:BD +1.0 mg T-2毒素/kg和0.2 mg Se/kg。
3.6 硒和T-2毒素对JAK-STAT信号介导的炎症和细胞凋亡的影响
在与JAK-STAT信号转导介导的脾脏炎症和凋亡相关的9个基因中,有6个基因受到日粮T-2毒素或Se的影响。与BD+Se组相比,BD和BD+T-2组增加(P<0.05)JAK 3、STAT1、STAT3、p53和JNK的mRNA水平,但降低BCL-2的mRNA水平。值得注意的是,日粮T-2毒素补充加重了缺硒组JAK3、STAT3、p53和JNK mRNA丰度的增加,而补硒缓解了这些变化。此外,T-2毒素和硒对脾脏中(IL-1β、IL-6和TNF-α的mRNA丰度没有影响(P>0.05)。然而,就这些基因的mRNA水平而言,没有观察到Se和T-2毒素之间的相互作用。
WB结果显示,脾组织中Jak3,P-Stat1,P53,STAT1,IL-6,Cleaved-Caspase 3,Bax,Bax-XL和Bcl-2的蛋白表达均受到影响日粮T-2毒素、硒及其交互作用的影响(P<0.05)(图6B和C),其中,T-2毒素对脾脏IL-6、STAT1、p53、Cleaved-Caspase 3、Bax,Bcl-2和Bcl-XL的影响最大(P<0.05 ),其中4种(包括IL-6、STAT1、cleaved-caspase 3和BCL-2和JAK3受硒的影响。值得注意的是,日粮中添加硒减轻了(P<0.05)T-2毒素引起的JAK3、STAT1、cleaved-caspase 3和BCL-2的变化。脾脏STAT1、cleaved-caspase 3和BCL-2表达受到T-2毒素和硒之间的相互作用影响(P<0.05)。
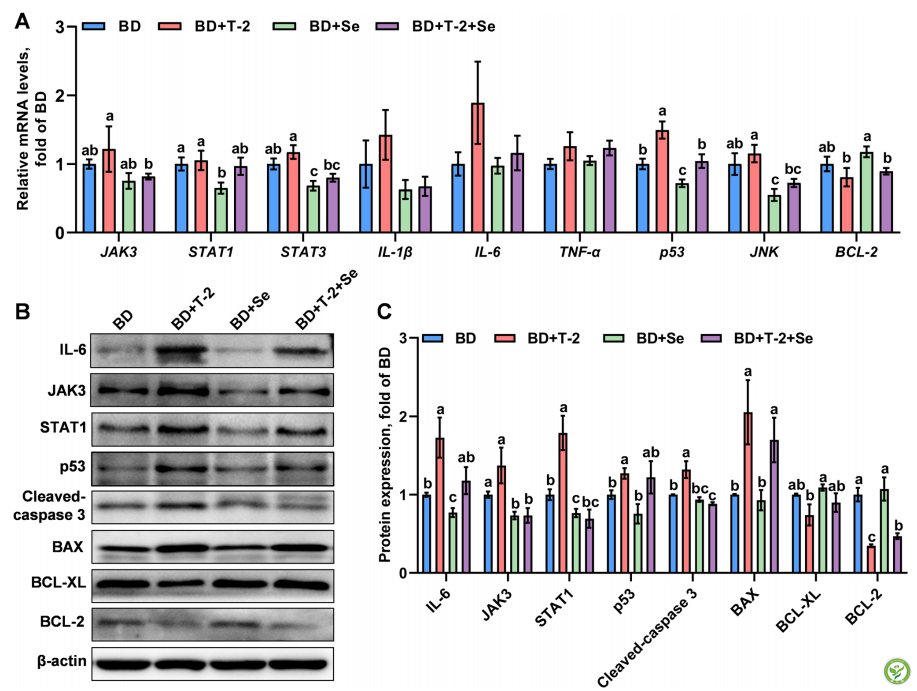
图6:日粮硒水平和T-2毒素对鸡脾脏中JAK-STAT信号通路介导的炎症和凋亡相关基因表达的影响。A JAK-STAT信号通路相关基因的相对mRNA水平。B JAK-STAT信号通路相关蛋白表达。C. JAK-STAT信号通路中涉及的蛋白质的相对丰度。BD:基础饮食; BD+T-2:BD加1.0 mg T-2毒素/kg; BD+Se:BD + T-2+Se:BD+T-2毒素1.0 mg/kg + Se 0.2 mg/kg。
4、结论
T-2毒素≥1.0mg/kg损害肉鸡生长性能,诱导免疫器官损伤,并可引起组织病理学改变和氧化还原失衡。目前的研究基于对肠道微生物群-脾脏轴的了,解释了T-2毒素对脾脏的损伤。联合微生物学和转录组学分析结果显示,T-2毒素主要通过促进IL-6/JAK/STAT 1信号介导的炎症和凋亡破坏肠道微生物群稳态和脾脏转录组。此外,本研究发现,日粮中添加硒可保护雏鸡免受T-2毒素诱导的不良反应,其机制可能与抑制IL-6/JAK/STAT 1信号通路的激活有关。这些发现系统地揭示了T-2毒素在鸡中潜在的免疫毒性,提示补硒有可能成为控制T-2毒素风险的治疗策略。
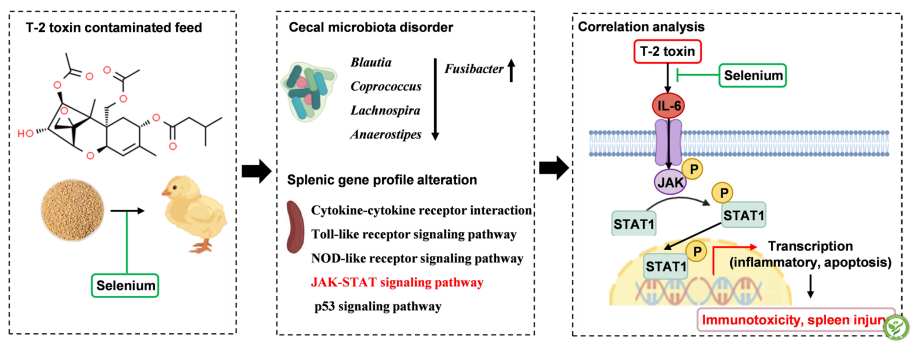
图7:T-2毒素对雏鸡的免疫毒性及硒通过肠道菌群-脾轴整合调控网络对T-2毒素的缓解作用示意图
1、摘要
T-2毒素是环境和食物链中普遍存在的毒性最大的单端孢菌毒素之一,给动物和人类带来严重的健康危害,并能显著损害免疫功能。本研究通过整合转录组和微生物组,全面解释了T-2毒素通过肠道微生物-脾脏轴对脾脏的影响。结果表明,日粮T-2毒素≥1.0 mg/kg显著抑制肉仔鸡的生长性能,引起脾脏损伤,并伴有氧化应激和组织病理学损伤。盲肠微生物组分析表明,T-2毒素暴露导致肠道微生物生态失调,特别是导致一些增强肠道屏障和减少炎症的有益菌属减少。转录组分析表明,T-2毒素暴露直接引起脾脏炎症和免疫相关信号,如细胞因子-细胞因子受体相互作用、Toll样受体信号通路、NOD样受体信号通路和JAK-STAT信号通路。
此外,通过联合转录组和微生物组分析,发现T-2毒素诱导的脾脏损伤与IL-6/JAK/STAT1信号通路介导的炎症和凋亡的异常激活有关,并通过western bolt分析验证。值得注意的是,日粮硒补充剂可以通过抑制IL-6/JAK/ STAT1信号相关基因的表达来保护雏鸡免受T-2毒素诱导的对生长性能和脾脏损伤的不利影响。总之,本研究为T-2毒素在鸡脾脏中的免疫毒性机制提供了新的见解,并强调了硒在减轻T-2毒素诱导的免疫毒性方面的潜力。
2、试验设计
3、结果与分析
3.1 T-2毒素降低生长性能并诱导脾损伤
4组肉鸡的初始体重没有差异(图1A)。经过2周的试验处理后,添加1.0mg T-2毒素/kg的日粮有降低(P=0.06)体重(12.4%)的趋势,添加3.0和6.0mg/kg的日粮有降低(P<0.05)最终体重(17.6–27.2%)的趋势(图1A)。在不同剂量下,饮食中T-2毒素补充≥1.0 mg/ kg可使体重增加减少13.7–30.2%(P< 0.05)(图1B)。此外,组织学结果显示,日粮补充T-2毒素诱导免疫组织损伤(图1C)。具体而言,与对照组相比,日粮添加T-2毒素≥1.0 mg/kg诱导脾淋巴细胞减少、网状上皮细胞增生和轻度充血,尤其是添加6.0 mg/kg T-2毒素的日粮诱导严重充血和巨噬细胞增生。同时,T-2毒素引起淋巴细胞坏死和耗竭、局灶性坏死、皮质淋巴细胞增殖、胸腺血管充血和法氏囊完整性的丧失。此外,T-2毒素诱导脾脏氧化损伤,与对照组相比,随着日粮T-2毒素剂量的增加,MDA和PC浓度分别增加55.7-97.3%和182.0-393.1%(P<0.05),T-AOC分别减少24.3-35.9%(P<0.05),呈剂量依赖性(图1D-F)。
3.2 T-2毒素对盲肠菌群稳态的影响
为更好地了解T-2毒素对盲肠细菌组成的影响,对对照组和实验1中6.0 mg/kg T-2毒素组肉鸡进行了16 S rRNA微生物基因的高通量测序(图2)。根据序列相似性(>97%)将符合条件的序列聚类为OTU,T-2毒素处理组中OTU数量较少,两组共有499个OTU(图2A)。基于分类学组成分析,T-2毒素改变了盲肠细菌在门和属水平上的相对丰度。所有来自盲肠食糜样品的合格序列归属于7个已知门。在所有样品中,以厚壁菌门、拟杆菌门、变形菌门和软壁菌门为优势菌门(图2B)。与对照组相比,添加6.0mg/kg的T-2毒素可显著降低厚壁菌门和软壁菌门的丰度,提高拟杆菌门和变形菌门的丰度;并从所有合格序列中鉴定出50个优势属,包括拟杆菌属、嗜双歧杆菌属、乳杆菌属、粪大肠杆菌属和示波螺旋菌属(图2C)。微生物区系的α多样性通过Chao 1分析评估物种丰富度,Shannon指数评估物种均匀度。与对照组相比,在日粮中添加6.0 mg T-2毒素/ kg不会影响Chao 1指数(图2D),但会降低Shannon指数(图2E)。为确定两组之间不同的细菌分类群,使用了LDA>3的LEfSe分析。结果在对照组和T-2毒素组的微生物群中分别鉴定了8个和2个区别性特征(图2F)。此外,菌属的丰度显示,与对照组相比,T-2毒素组中的Blautia、Coprococcus、Lachnospira、Coprobacillus、Anaerostipes 属的丰富度显著减少;然而,与对照组相比,在T-2毒素处理下,Fusibacter丰度显著增加(图2L)。
图2:日粮T-2毒素对盲肠微生物组的影响。A对照组和6.0 T-2组中OTU的数量。B门水平的肠道细菌分布。C属水平的肠道细菌分布。D和E T-2毒素对Chao 1指数的影响。E T-2毒素对Shannon指数的影响。F线性判别分析(LDA)效应大小(LEfSe)微生物群组成分析。G-L Blautia、Coprococcus、Lachnospira、Coprobacillus、Anaerostipes 、Fusibacter相对丰度。对照,基础饮食;6.0 T-2,基础日粮添加6.0 mg/kg T-2毒素。
3.3 T-2毒素对脾脏转录组的影响
为进一步揭示T-2毒素的毒性机制,对实验1的肉鸡脾脏组织进行了转录组分析。差异表达(DE)分析结果表明,T-2毒素处理的肉鸡脾脏中存在T-2毒素处理应答基因(图3)。与对照组相比,在日粮中补充1.0、3.0和6.0mg/kgT-2毒素时,分别有2489、1943和3936个基因表现出1.5倍或更高(P<0.05)的DE(图3A-C)。值得注意的是,在不同的T-2毒素剂量下,有907个基因具有相似的变化(图3D)。此外,GO和KEGG通路分析确定了共表达的DEG。主要GO类别的不同分布如图3E所示。在生物过程(BP)类别中,丰富了细胞周期、防御反应、免疫系统过程、对生物刺激的反应和对应激的反应。在细胞化合物(CC)类别中,染色质、染色体、胞外区细胞外空间和超分子复合物是GO富集。CCR趋化因子受体结合、CCR 6趋化因子受体结合、趋化因子活性、趋化因子受体结合和蛋白质异二聚体活性也是GO表达最多的类别。KEGG富集结果表明,KEGG的主要通路包括细胞因子-细胞因子受体相互作用、Toll样受体信号通路、核苷酸寡聚化结构域(NOD)样受体信号通路、IL-17信号通路、细胞坏死、凋亡-多物种、细胞因子生长因子和Janus激酶(JAK)-信号传导子和转录激活子(STAT)信号通路(图3F)。
图3 :T-2毒素对脾脏转录组的影响。A-C火山图分别显示1.0 T-2、3.0 T-2、6.0 T-2毒素与对照中的DEG。D维恩图分析1.0 T-2毒素与对照、3.0 T-2毒素与对照、6.0 T-2毒素相对于对照DEG数量。E GO功能富集分析。F KEGG途径富集分析,对照,基础饲料; 1.0 T-2、3.0 T-2和6.0 T-2,基础饲料添加1.0、3.0和6.0 mg/kg T-2毒素。
3.4 转录组和微生物组联合分析揭示T-2毒素对脾脏的毒性机制
DEG与最丰富的细菌属的相关性分析表明,有17个DEG与10个属的DEG具有显著的相关性(图4A)。其中,Faecalibacterium_prausnitzii和Ruminococcus与大多数基因相互作用最高。此外,在KEGG途径分析中具有显著相关性的这些DEG的注释途径主要与炎症反应和免疫调节有关,包括ECM-受体相互作用、JAK-STAT信号传导途径和细胞因子-细胞因子受体相互作用(图4B)。基于筛选的KEGG富集途径,利用Western blot分析JAK-STAT信号通路在T-2毒素对肉鸡脾脏的影响中的作用(图4C和D)。值得注意的是,与对照相比,日粮补充1.0、3.0和6.0 mg T-2毒素/kg均增加了(P<0.05)JAK 2、JAK 3和c-Jun-N-末端激酶(JNK)的蛋白生成。对照组相比,3.0和6.0 mg T-2毒素/kg组显著增加(P<0.05)STAT 1、IL-6和p53蛋白的表达。有趣的是,6.0 mg T-2毒素/kg组的脾中p-STAT 1表达比其他三组低。然而,BCL-2的蛋白质产生不受日粮T-2毒素剂量的影响(P>0.05)(图4C和D)。
图4:转录组与微生物组的联合分析。A属与DEG之间的Pearson相关性分析。B KEGG途径富集分析DEG与属显著相关性。C参与JAK-STAT信号通路的蛋白相对丰度。D参与JAK-STAT信号通路的蛋白的相对丰度。
3.5 硒和T-2毒素对生长性能、脾脏氧化还原状态和组织病理学的影响
四组肉鸡的初始体重差异不显著(图5A)。在3周的实验处理后,与BD组相比,日粮T-2毒素补充降低了(P<0.05)终末体重(20.1%),体增重(21.9%),采食量(13.7%)和增重/饲料(9.2%),而补硒(添加硒为单质纳米硒,由北京德元顺生物科技有限公司提供)减轻了T-2毒素引起的这些变化(图5B-E)。与BD组相比,BD +T-2组增加了(P<0.05)GPX活性,BD+Se+T-2组降低(P<0.05)MDA含量。且与BD+T-2组相比,BD+Se+T-2组GPX活性升高(P<0.05)。然而,TrxR活性不受日粮Se缺乏和T-2毒素补充的显著影响(图5H)。除GPX活性外,没有观察到Se和T-2毒素之间的相互作用。此外,脾组织学结果显示,日粮T-2毒素诱导的脾损伤,这是由淋巴细胞数量减少和组织细胞增殖的减少所表明(图5I)。单独硒缺乏也可引起脾淋巴细胞减少和组织细胞增殖能力下降,而BD+Se组未见明显病理改变。
图5:日粮硒水平和T-2毒素对生长性能(A-E)、脾脏氧化还原状态(F-H)和组织病理学(I)的影响。BD:基础日粮; BD+T-2:BD +1.0 mg T-2毒素/kg; BD+Se,BD +0.2 mg Se/kg; BD+T-2+Se:BD +1.0 mg T-2毒素/kg和0.2 mg Se/kg。
3.6 硒和T-2毒素对JAK-STAT信号介导的炎症和细胞凋亡的影响
在与JAK-STAT信号转导介导的脾脏炎症和凋亡相关的9个基因中,有6个基因受到日粮T-2毒素或Se的影响。与BD+Se组相比,BD和BD+T-2组增加(P<0.05)JAK 3、STAT1、STAT3、p53和JNK的mRNA水平,但降低BCL-2的mRNA水平。值得注意的是,日粮T-2毒素补充加重了缺硒组JAK3、STAT3、p53和JNK mRNA丰度的增加,而补硒缓解了这些变化。此外,T-2毒素和硒对脾脏中(IL-1β、IL-6和TNF-α的mRNA丰度没有影响(P>0.05)。然而,就这些基因的mRNA水平而言,没有观察到Se和T-2毒素之间的相互作用。
WB结果显示,脾组织中Jak3,P-Stat1,P53,STAT1,IL-6,Cleaved-Caspase 3,Bax,Bax-XL和Bcl-2的蛋白表达均受到影响日粮T-2毒素、硒及其交互作用的影响(P<0.05)(图6B和C),其中,T-2毒素对脾脏IL-6、STAT1、p53、Cleaved-Caspase 3、Bax,Bcl-2和Bcl-XL的影响最大(P<0.05 ),其中4种(包括IL-6、STAT1、cleaved-caspase 3和BCL-2和JAK3受硒的影响。值得注意的是,日粮中添加硒减轻了(P<0.05)T-2毒素引起的JAK3、STAT1、cleaved-caspase 3和BCL-2的变化。脾脏STAT1、cleaved-caspase 3和BCL-2表达受到T-2毒素和硒之间的相互作用影响(P<0.05)。
图6:日粮硒水平和T-2毒素对鸡脾脏中JAK-STAT信号通路介导的炎症和凋亡相关基因表达的影响。A JAK-STAT信号通路相关基因的相对mRNA水平。B JAK-STAT信号通路相关蛋白表达。C. JAK-STAT信号通路中涉及的蛋白质的相对丰度。BD:基础饮食; BD+T-2:BD加1.0 mg T-2毒素/kg; BD+Se:BD + T-2+Se:BD+T-2毒素1.0 mg/kg + Se 0.2 mg/kg。
4、结论
T-2毒素≥1.0mg/kg损害肉鸡生长性能,诱导免疫器官损伤,并可引起组织病理学改变和氧化还原失衡。目前的研究基于对肠道微生物群-脾脏轴的了,解释了T-2毒素对脾脏的损伤。联合微生物学和转录组学分析结果显示,T-2毒素主要通过促进IL-6/JAK/STAT 1信号介导的炎症和凋亡破坏肠道微生物群稳态和脾脏转录组。此外,本研究发现,日粮中添加硒可保护雏鸡免受T-2毒素诱导的不良反应,其机制可能与抑制IL-6/JAK/STAT 1信号通路的激活有关。这些发现系统地揭示了T-2毒素在鸡中潜在的免疫毒性,提示补硒有可能成为控制T-2毒素风险的治疗策略。
图7:T-2毒素对雏鸡的免疫毒性及硒通过肠道菌群-脾轴整合调控网络对T-2毒素的缓解作用示意图
 电话: 010-80854808
电话: 010-80854808
 传真:010-89525360
传真:010-89525360
 邮箱:dysbio@163.com
邮箱:dysbio@163.com
